I
Ко́рень (radix)
один из основных вегетативных органов листостебельных растений (за исключением мхов), служащий для прикрепления к субстрату, поглощения из него воды и питательных веществ, первичного превращения ряда поглощаемых веществ, синтеза органических соединений, дальнейшего перемещения их в др. органы растения, а также для выделения некоторых продуктов обмена. У некоторых растений К. несут дополнительные функции, например вместилища запасных веществ, а у корнеотпрысковых растений (См.
Корнеотпрысковые растения) - органа вегетативного размножения. К. - осевой орган растения, по происхождению родственный стеблю: у предков наземных растений при выходе их на сушу на нижнем конце
Спорофита образовались корневищеподобные веточки - прототип К. Морфологически К. отличается от стебля отсутствием листьев, наличием чехлика и эндогенным ветвлением (возникновением боковых К. во внутренних частях К. - в
Перицикле)
. Типичный К. имеет узкоцилиндрическую или нитевидную форму. Зачаток К. существует уже в зародыше семени, при прорастании которого он развивается в главный К. Для К. характерно акропетальное ветвление (более молодые боковые К. и их зачатки появляются ближе к верхушке К.). У многих растений, кроме главного и боковых К., имеются придаточные. Внешне они не отличаются от боковых и несут те же функции, но обычно развиваются на др. органах растения - стеблях, листьях, подземных и надземных видоизменённых побегах (луковицах, клубнях, корневищах и т. д.), а также на старых К. Образование придаточных К. определяет возможность вегетативного размножения растений. Кончик К. (
рис. 1) прикрыт корневым чехликом (См.
Корневой чехлик)
, защищающим его от повреждений. К. растет в длину за счёт деления клеток верхушечной (апикальной) меристемы (См.
Меристема)
, в т. н. зоне деления, и значительного роста их в длину в зоне роста, или растяжения, длина которой обычно равна 1-2
мм (иногда до 10
мм)
. За зоной роста расположена зона поглощения и дифференциации; самый поверхностный слой клеток этой зоны дифференцируется в эпиблему (См.
Эпиблема)
, клетки которой образуют
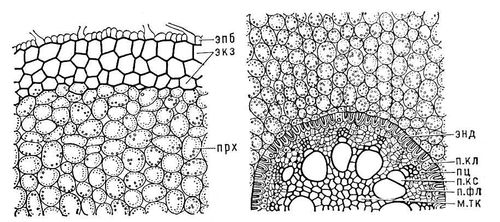
Корневые волоски, увеличивающие поверхность поглощения К. в 5-20 раз. Дифференциация меристемы в первичные постоянные ткани начинается в зоне роста, но отчётливо выявляется в зоне поглощения и в т. н. зоне проведения. Первичное анатомическое строение К. всех семенных растений сходно (
рис. 2). Под слоем клеток эпиблемы расположена многослойная первичная кора, которая состоит из тонкостенных живых паренхимных клеток. У некоторых растений оболочки 1-3 наружных слоев клеток коры (
Экзодерма) опробковевают и по отмирании эпиблемы берут на себя защитную функцию. Самый внутренний слой первичной коры (
Эндодерма) состоит из одного слоя клеток, оболочки которых частично опробковевают или одревесневают и нередко утолщаются; лишь некоторые клетки (т. н. пропускные) остаются мало измененными. Эндодерма окружает центральный цилиндр, или стель, К. Один или несколько наружных слоев его клеток образуют перицикл, тонкостенные живые клетки которого долго сохраняют меристематическую активность; в нём закладываются боковые К., придаточные почки; при его участии осуществляется вторичное утолщение К. Большую часть центрального цилиндра занимает сложный проводящий радиальный пучок, в котором чередуются элементы ксилемы (См.
Ксилема)
и флоэмы (См.
Флоэма)
. У однодольных растений первичное анатомическое строение К. сохраняется всю жизнь, у двудольных и голосеменных оно сменяется вторичным (
рис. 3). При этом в центральном цилиндре пучковый камбий откладывает (между флоэмой и ксилемой) к центру вторичную ксилему (древесину), а к периферии - вторичную флоэму (луб). Из межпучкового камбия (См.
Камбий)
, находящегося против лучей первичной ксилемы в перицикле, образуется паренхима первичного сердцевинного (лубо-древесинного) луча, а пробковый камбий (феллоген) откладывает кнаружи клетки пробки; она становится покровной тканью вторично утолщённого К., а вся первичная кора сбрасывается. Эти изменения обусловливают разрастание К. в толщину. Различают т. н. ростовые К., которые быстро растут (в благоприятных условиях - в среднем от 1 до 3
см в сутки), утолщаются, рано опробковевают, составляют скелет корневой системы и в ряде случаев обеспечивают вегетативное размножение растения, и "сосущие" К. - тонкие, нежные, короткие, медленно растущие, большей частью недолговечные.
Совокупность К. одного растения называется корневой системой, общую форму и характер которой определяет соотношение роста главного, боковых и придаточных К. (
рис. 4). При преобладающем росте главного К. образуется стержневая корневая система (люпин, хлопчатник); при слабом росте или раннем отмирании главного и преобладающем развитии большого числа придаточных К. образуется мочковатая корневая система (однодольные, из двудольных - виды лютика, подорожника и др.). К. у хлебных злаков (ржи, пшеницы) проникают на глубину 1-1,5
м, у люцерны -до
10 м, у древесных растений могут углубляться на 10-12
м, хотя обычно твёрдый грунт препятствует проникновению их глубже 3-5
м; площадь, занятая корневой системой одного растения хлебных злаков, достигает в поперечнике 40-60
см, тыквы - 6-8
м, у древесных растений обычно в несколько раз превышает диаметр кроны, достигая 10-18
м. Суммарная длина К. однолетнего растения может составлять несколько километров, а общая поверхность К., учитывая поверхность волосков, во много раз превышает поверхность надземных органов. По расположению в почве в зависимости от среды обитания различают корневые системы специализированные (поверхностные или глубинные) и универсальные, равномерно развивающиеся вширь и вглубь. Например, в северной лесной зоне на подзолистых почвах, часто пересыщенных влагой, плохо аэрируемых, с трудно проницаемой подпочвой, корневая система растений на 90-95\% сосредоточена в поверхностных слоях (10-15
см)
. В зоне полупустынь и пустынь у одних растений имеются только поверхностные К., которые используют осадки ранней весны (
Эфемеры) либо влагу дождей и конденсационную влагу, оседающую в верхних слоях почвы в ночное время (кактусы); у других К. достигают грунтовых вод (у верблюжьей колючки). Универсальные корневые системы используют в разное время влагу разных горизонтов, вследствие чего растения могут вегетировать всё лето (джузгун, саксаул, эфедра). В зонах недостаточного увлажнения ярко проявляется ярусность в расположении К. различных видов растений, произрастающих совместно на одной площади. Соотношение между К. и надземными частями растений также зависит от почвенно-климатических условий. В северной влажной зоне масса корневой системы в 5-10 раз уступает массе надземных частей. Через К. растения поглощают из почвы главным образом ионы минеральных солей, а также некоторые продукты жизнедеятельности почвенных микроорганизмов и корневые выделения др. растений. Поглощённые К. соединения азота, фосфора и серы взаимодействуют с притекающими из листьев продуктами фотосинтеза с образованием аминокислот, нуклеотидов и др. органических соединений. По сосудам ксилемы элементы в форме ионов (калий, кальций, магний, фосфор) или органических молекул (азот, сера) в результате действия корневого давления (См.
Корневое давление) и транспирации (См.
Транспирация) передвигаются в листья и стебли. В К. синтезируются также алкалоиды (например, никотин), гормоны роста (
Кинины, Гиббереллины) и др. физиологически активные вещества. У некоторых растений, преимущественно семейство бобовых, в К. поселяются бактерии, вызывающие образование клубеньков (из паренхимы К.). К. растений выделяют в почву ионы минеральных солей, аминокислоты, сахара, мочевину, фенольные соединения и др. У одних растений (например, семейство паслёновых) К. выделяют также
Ауксины и вещества типа гиббереллинов, стимулирующие рост растений, у др. - студнеобразные вещества, образующие корнезащитные чехлы. Выделения К. способствуют развитию в ризосфере почвенных микроорганизмов. С возрастом растения количество выделений и численность ризосферных микроорганизмов уменьшаются. Клубеньковые бактерии играют важную роль в азотном питании растений и повышении плодородия почвы. На поверхности или внутри К. многих древесных и травянистых растений поселяются грибы, образуя т. н. микоризу (См.
Микориза)
; в таких случаях корневые волоски на К. не развиваются.
На К. многих растений образуются придаточные почки, дающие надземные побеги (у корнеотпрысковых растений), У ряда растений К. служат местом отложения запасных питательных веществ (см.
Корнеплоды). У некоторых деревьев тропических лесов близ основания стволов или от ветвей отходят боковые или придаточные К. - досковидные, ходульные, столбовидные, служащие для опоры и питания. У лазящих лиан (См.
Лианы) (например, у плюща) развиваются корни-прицепки. В тропиках многие
Эпифиты образуют придаточные воздушные К., поглощающие через многослойную поверхностную ткань (т. н. веламен) воду из атмосферных осадков. У некоторых тропических эпифитных орхидей, а также у растений семейства подостемоновых листья и стебли недоразвиты и их вегетативные органы представлены главным образом плоскими зелёными К., выполняющими функции ассимиляции. У растений, живущих на бедных кислородом почвах (например, у болотного кипариса, у мангровых деревьев и др.), имеются
дыхательные К., или пневматофоры, верхушки которых расположены над почвой или водой и снабжают подземные органы воздухом. У некоторых пальм и растений семейства мареновых часть горизонтальных К. превращается в защитные колючки. К. паразитирующих на деревьях растений, например омелы, имеют вид длинных цилиндрических тяжей, расположенных в коре дерева. У растений-паразитов (заразиха, повилика и др.) и полупаразитов (марьянник, погремок и др.) корневая система развивается слабо; при этом окончания некоторых К. внедряются в виде особых присосок (
гаусторий) в тело растения-хозяина, высасывая из него питательные вещества. У некоторых растений (например, у роголистника, пузырчатки и др.) К. отсутствуют, что связано со специфическими условиями их существования. О функции К. см. также
Водный режим растений, Минеральное питание растений. К. многих растений широко используются человеком. Они имеют большое пищевое и хозяйственное значение. К., содержащие крахмал, сахара, масла, алкалоиды, гуттаперчу, красящие и др. ценные вещества, применяются в медицине и промышленности. Растения с мощно развитой корневой системой используются для закрепления подвижных песков, оврагов и эродированных почв.
Лит.: Красовская И. В., Обзор работ по морфологии и физиологии корней, "Тр. по прикладной ботанике генетике и селекции", 1928, т. 18, в. 5; её же, Закономерности строения корневой системы хлебных злаков, "Ботанический журнал", 1950, т. 35, № 4; Шалыт М. С., Подземная часть некоторых луговых, степных и пустынных растений и фитоценозов, "Труды Ботанического института АН СССР. Сер. 3, Геоботаника", 1950, в. 6; Сабинин Д. А., Физиологические основы питания растений, М., 1955; Качинский Н. А., Почва, её свойства и жизнь, М., 1956; Колесников В. А., Корневая система плодовых и ягодных растений и методы сё изучения, М., 1962; Федоров А. А., Кирпичников М. Э. и Артюшенко З. Т., Атлас по описательной морфологии высших растений, [т. 2], М.- Л., 1962; Колосов И. И., Поглотительная деятельность корневых систем растений, М., 1962; Рахтеенко И. Н., Рост и взаимодействие корневых систем древесных растений, Минск, 1963; Воронин Н. С., Эволюция первичных структур в корнях растений, "Уч. зап. Калужского педагогического института", 1964, в. 13; Бойко Л. А., физиология корневой системы растений в условиях засоления, Л., 1969; Эсау К., Анатомия растений, пер, с англ., М., 1969.
О. Н. Чистякова, Р. П. Барыкина, Д. Б. Вахмистров.
Рис. 1. Молодой корень проростка пшеницы: А - схема строения: к.чх - корневой чехлик, кл - калиптроген, з.д - зона деления, з.р - зона растяжения (роста), з.в - зона всасывания, з.п - зона проведения; Б - периферические клетки отдельных зон при большом увеличении.
Рис. 2. Поперечный срез корня касатика в зоне проведения: слева - эпб - эпиблема, экз - экзодерма, прх - запасающая паренхима первичной коры; справа - энд - эндодерма, пкл - пропускная клетка, пц - перицикл, пкс - первичная ксилема, п. фл - первичная флоэма, м. тк - механическая ткань.
Рис. 3. Схема вторичного утолщения корня: А - первичное строение; Б, В - вторичное утолщение; эпб - эпиблема, п. к. - первичная кора, энд - эндодерма, пц - перицикл, п. фл. - первичная флоэма, кмб - камбий, п. кс. - первичная ксилема, вт. фл. - вторичная флоэма, вт. кс. - вторичная ксилема, флг - феллоген, прд - перидерма, разр. энд. - разрушенная эндодерма, п. с. л. - первичный сердцевинный луч, в. с. л. - вторичный сердцевинный луч.
Рис. 4. Различные формы корней и типы корневой системы: 1, 4 - стержневая; 5 - мочковатая; 2, 3 - корнеплоды (моркови и свёклы).
II
Ко́рень
в
математике, 1) К. степени
n из числа
а - число
х (обозначаемое
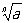
),
n-я степень которого равна
а (то есть
xn = а)
. Действие нахождения К. называют извлечением корня (См.
Извлечение корня)
. При
а ≠ 0 существует
n различных значений К. (вообще говоря, комплексных); например, значениями
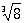
являются: 2; -1+i√3; -1-i√3. К нахождению К. из чисел приводили различные геометрические задачи математиков глубокой древности. Среди вавилонских клинописных текстов (2-е тысячелетие до н. э.) имеются описания приближённого нахождения квадратного К. и таблицы квадратных К., а в египетских папирусах встречается для действия извлечения К. и особый знак. Древнегреческие математики установили несоизмеримость стороны квадрата с его диагональю (равной
а√2, если
а - сторона), что позднее привело к открытию иррациональности.
Ариабхата (5 в.) дал правила для извлечения квадратных и кубических К.
Омар Хайям (2-я половина 11 - начало 12 вв.), аль
-Каши (15 в.), немецкий математик М. Штифель (16 в.) извлекали К. высших степеней, исходя из формулы для (
а+b)
n. Л.
Эйлер (18 в.) дал сохранившие своё значение до наших дней приближённые способы извлечения К. Квадратные К из отрицательных чисел, встречающиеся в 16 в. у Дж. Кардана (См.
Кардано) и Р.
Бомбелли, привели к открытию комплексных чисел.
2) К. алгебраического уравнения
a0xn + a1xn-1+... + an-1x + an = 0 (1)
- число с, которое после подстановки его вместо х обращает уравнение в тождество. К. уравнения (1) называется также и К. многочлена
f (x) = a0xn + a1xn-1+... + an-1x + an.
Если
с является К. многочлена
f (x), то
f (x) делится без остатка на
х-с. См. также
Многочлен, Уравнение. слова, морфема (морфа), являющаяся общим элементом группы производных от него слов, имеющих в основе общее лексическое значение. К. выявляется путём устранения всех аффиксов как словоизменительных, так и словообразовательных, например русский "луч" в словах "луч", "лучи", "лучевой", "излучение". В некоторых языках К. может подвергаться звуковым изменениям (см.,
Внутренняя флексия)
, например немецкий find/fand/fund в словах finden, fand, gefunden - "находить"; арабский h-s-b в словах hisāb - "счёт", hāsib - "счётчик", muhtasib - "koнтролёр мер и весов". Сложные слова могут содержать два и более К., например "лучевидный". Этимологический К. (см.
Этимон)
- форма и значение слова, мыслимые как исходные. Например "кол" со значением "круг" в словах "колесо", "около".